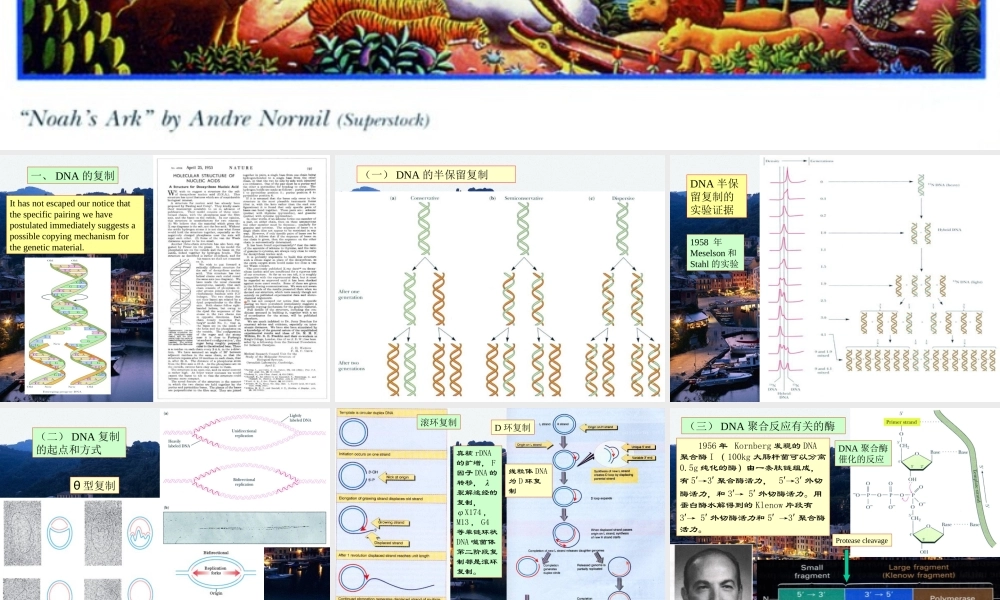
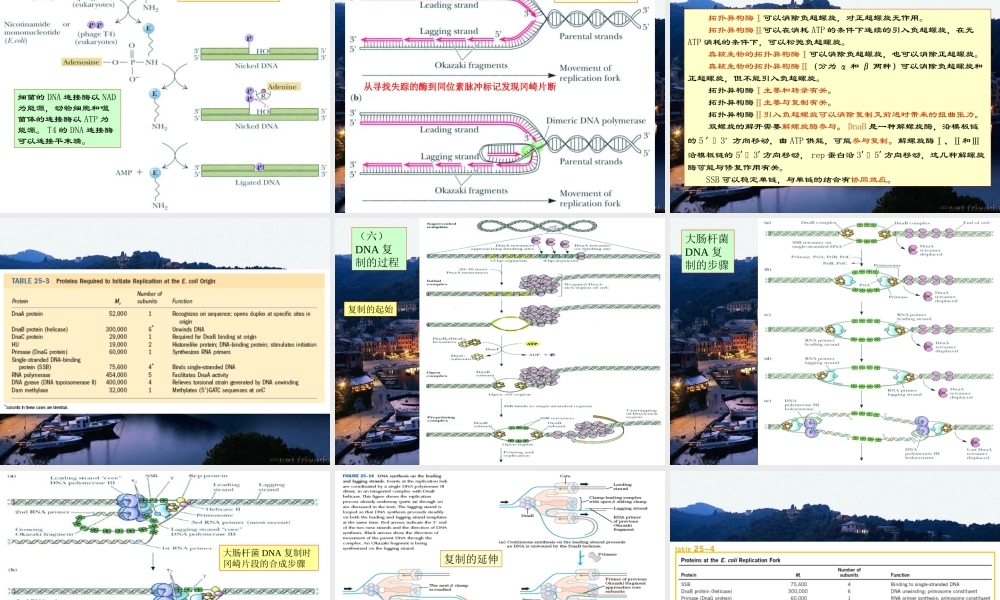
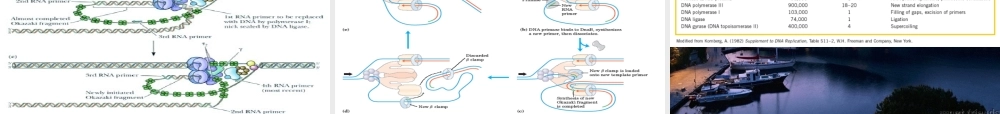
DNA的复制和修复第34章*Ithasnotescapedournoticethatthespecificpairingwehavepostulatedimmediatelysuggestsapossiblecopyingmechanismforthegeneticmaterial.一、DNA的复制(一)DNA的半保留复制DNA半保留复制的实验证据1958年Meselson和Stahl的实验(二)DNA复制的起点和方式θ型复制D环复制滚环复制真核rDNA的扩增,F因子DNA的转移,λ裂解途经的复制,φX174,M13,G4等单链环状DNA噬菌体第二阶段复制都是滚环复制。线粒体DNA为D环复制1956年Kornberg发现的DNA聚合酶I(100kg大肠杆菌可以分离0.5g纯化的酶)由一条肽链组成,有5′→3′聚合酶活力,5′→3′外切酶活力,和3′→5′外切酶活力。用蛋白酶水解得到的Klenow片段有3′→5′外切酶活力和5′→3′聚合酶活力。(三)DNA聚合反应有关的酶DNA聚合酶催化的反应ProteasecleavageDNApolymeraseIhasthreeenzymaticactivitiesinasinglepolypeptidechain,whichcanbecleavedintotwofunctionalpartsbymildproteasetreatment.图示为Klenow片段与DNA的结合,模板链(12nt)为蓝色,引物链(14nt)为红色。DNA聚合酶I的Klenow片段与DNA的相互作用DNA聚合酶I的5′3′外切酶可以从单链切口的5′末端切除核苷酸,5′3′聚合酶可以用脱氧核苷酸填补缺口。DNA聚合酶I的校对作用DNA聚合酶的结构DNA聚合酶Ⅲ的β亚基二聚体与DNA结合的空间关系(顶部观)。DNA聚合酶Ⅲ的β亚基二聚体与DNA结合的空间关系(立体图)。DNA聚合酶Ⅱ有不少于7个亚基,可能参与DNA的修复,1999年发现的DNA聚合酶Ⅳ和Ⅴ可以复制有较多损伤的DNA。DNA连接酶的作用机制细菌的DNA连接酶以NAD为能源,动物细胞和噬菌体的连接酶以ATP为能源。T4的DNA连接酶可以连接平末端。(四)DNA的半不连续复制从寻找失踪的酶到同位素脉冲标记发现冈崎片断(五)DNA复制的拓扑性质拓扑异构酶Ⅰ可以消除负超螺旋,对正超螺旋无作用。拓扑异构酶Ⅱ可以在消耗ATP的条件下连续的引入负超螺旋,在无ATP消耗的条件下,可以松弛负超螺旋。真核生物的拓扑异构酶Ⅰ可以消除负超螺旋,也可以消除正超螺旋。真核生物的拓扑异构酶Ⅱ(分为α和β两种)可以消除负超螺旋和正超螺旋,但不能引入负超螺旋。拓扑异构酶Ⅰ主要和转录有关。拓扑异构酶Ⅱ主要与复制有关。拓扑异构酶Ⅱ引入负超螺旋可以消除复制叉前进时带来的扭曲张力。双螺旋的解开需要解螺旋酶参与。DnaB是一种解螺旋酶,沿模板链的5′3′方向移动,由ATP供能,可能参与复制。解螺旋酶Ⅰ、Ⅱ和Ⅲ沿...